Podać definicję pojęcia „gen” nie jest łatwo
W poprzednim paragrafie rozpatrzyliśmy, jak informacja o budowie DNA kolejno realizuje się w procesach transkrypcji, a potem translacji. Najpierw RNA-polimeraza syntezuje matrycowy RNA na cząsteczce DNA, a potem rybosom syntezuje białko, odczytując informację z matrycowego RNA. U eukariotów jeden matrycowy RNA zwykle zawiera informację o budowie całego typu cząsteczek białkowych. Ten matrycowy RNA syntezuje się z osobliwego odcinka DNA, który nazywa się genem.
Otóż informacja o kolejności reszt aminokwasowych w cząsteczce białka zakodowana jest w osobliwym odcinku cząsteczki DNA - genie. Jednak geny kodują nie tylko matrycowy RNA. Transportujące RNA, rybosomowe RNA i inne rodzaje RNA też są kodowane swymi genami, jednak te RNA nigdy nie ulegają translacji, zostając na zawsze RNA. Możemy więc sformułować szerszą definicję genu: gen - to odcinek DNA, który koduje cząsteczkę RNA1.
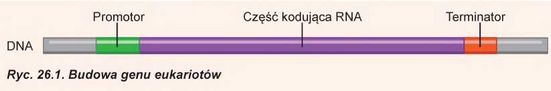
Rozpatrzmy budowę zwykłego genu, który koduje matrycowy RNA komórki zwierzęcej (ryc. 26.1). Gen, jak już wiemy, jest odcinkiem dwułańcuchowego DNA i tylko jeden łańcuch koduje mRNA. Ten łańcuch nazywa się łańcuchem antytreściowym (nić sensowna). Kolejność nukleotydów w nim jest komplementarna do matrycowego RNA, właśnie z tym łańcuchem będzie pracować RNA-polimeraza. Kolejność nukleotydów innego łańcucha DNA zbiega się z kolejnością nukleotydów matrycowego RNA kodującego się genem. Ten łańcuch nazywa się łańcuchem treściowym (nić niesensowna). Przy tym jeden łańcuch DNA może być antytreściowy dla jednych genów i treściowy - dla innych.

Teraz rozpatrzmy, jak rozmieszczają się różne odcinki genu wzdłuż cząsteczki DNA. Gen zaczyna się z odcinka, który odpowiada za przyłączenie RNA-polimerazy i początek translacji - promotora. Następnie za promotorem rozmieszczony jest odcinek genu, który koduje matrycowy RNA-część kodująca genu. Jak pamiętasz, matrycowy RNA zawiera start-kodon. Taki sam start-kodon zawiera gen. Za nim rozmieszczona
1 Jednak nawet taka definicja nie jest doskonała. Do niej nie pasują na przykład geny wirusów z RNA-genomem. Wiadomo, że często u eukariotów z jednego genu syntezuje się kilka różnych cząsteczek RNA. U prokariotów kilka genów może kodować jedną cząsteczkę RNA, o czym będzie mowa w tym paragrafie. Chociaż nasza definicja pojęcia „gen” jest dość ścisła, ona ukazuje jego istotę.
kolejność nukleotydów, kodujących aminokwasy w białku. W końcu kodujący odcinek kończy się stop-kodonem1. Jednak nawet po stop-kodonie transkrypcja powinna jakiś czas być kontynuowana: cząsteczka matrycowego RNA zawiera długi odcinek, który nie transluje się rybosomem, lecz jest ważny dla funkcjonowania matrycowego RNA. Gen kończy się odcinkiem, który odpowiada za ustanie transkrypcji - terminatorem. Terminator zapewnia odłączenie RNA-polimerazy od DNA i ustanie transkrypcji. Jak i w przypadku promotora, terminator może być po różnemu zorganizowany w różnych genach. Ciekawa rzecz, że kolejność, która wykonuje rolę terminatora w genach matrycowego RNAeukariotów, nie kończy transkrypcji natychmiast: RNA-polimeraza odczytuje jeszcze 100-200 nukleotydów i tylko wtedy „odpada” od DNA. Zbyteczne nukleotydy potem są odcinane enzymami komórki.
Geny prokariotów i eukariotów różnią się pod względem budowy
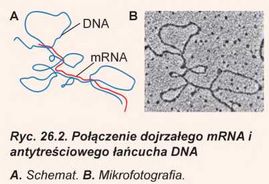
Jak już zaznaczaliśmy, jeden łańcuch DNA, a dokładniej - antytreściowy, jest całkowicie komplementarny do cząsteczki RNA syntezowanej na nim RNA-polimerazą. To znaczy, że jeżeli w probówce dodamy do tego łańcucha syntezowany na nim RNA, to one utworzą dwułańcuchową strukturę podobną do tej, którą tworzą dwa łańcuchy DNA. Natomiast gdy dodamy antytreściowy łańcuch DNA do dojrzałego matrycowego RNA kodowanego tym genem, to okaże się, że ilość komplementarnych odcinków nie będzie tak duża. Większa część antytreściowego DNA nie będzie komplementarna do matrycowego RNA i będzie tworzyć pętle, część komplementarna będzie w postaci kilku rozróżnionych odcinków (ryc. 26.2).
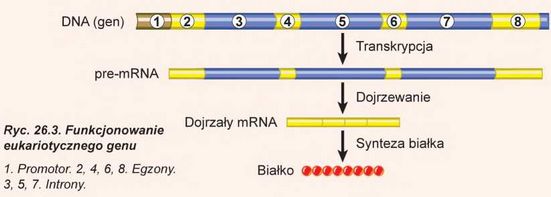
Tłumaczy się to tym, że u eukariotów po syntezie poprzednika matrycowego RNA z niego wycinane są nieduże odcinki. Te odcinki nazywają się intronami. Introny przeważnie nie kodują kolejności aminokwasowej białka. One bezpowrotnie usuwają się z cząsteczki RNA podczas jej dojrzewania. Przy tym odcinki kodujące, które pozostały - egzo-ny - łączą się między sobą. Proces wycinania intronów zachodzi w jądrze przy udziale małych jądrowych RNA. Ten proces jest nie- odjemną częścią dojrzewania matrycowego RNA. Czasami introny stanowią 90% całej kolejności genu. Wynika stąd, że w toku dojrzewania pre-mRNA skraca się dziesięciokrotnie. Funkcje intronów poznane są nie do końca. One są uważane za potomków szkodliwych kolejności DNA, które infekowały genom i bez kontroli namnożyły się w nim (tak zwany egoistyczny DNA). Według tej teorii wycinanie intronów - to proces oczyszczenia genów od niekodowanych kolejności, które ich zaraziły. Jednak komórki nauczyły się wykorzystywać introny do kodowania kilku białek jednym genem, zwiększając w ten sposób różnorodność własnych białek. Dlatego czasami introny kodują odcinki aminokwasowej kolejności białka. Przypuśćmy, że w procesie wycinania jeden intron pozostanie. Wtedy informacja, którą on zawiera, rybosom wykorzysta do budowy cząsteczki białka. Ciekawa rzecz, że wskutek tego procesu z jednego genu powstają różne matry-

cowe RNA, a z nich - różne białka. Przykładem takiego genu może być gen fibronektyny. On zawiera fragment, kodujący odcinek RNA, który u większości komórek nie wycina się, czyli rozpoznaje się jako egzon. Ten odcinek odpowiada za wiązanie z białkami międzykomórkowej substancji i jej włączenie do cząsteczki białka prowadzi do powstania nierozpuszczalnej formy fibronektyny. Jednak w komórkach wątroby ten odcinek rozpoznaje się jako intron i wycina się. Prowadzi to do kształtowania cyrkulującej formy fibronektyny rozpuszczonej w osoczu krwi. Otóż jeden gen koduje dwa białka. Uważa się, że u człowieka tak funkcjonuje większość genów, dlatego ilość białek przewyższa ilość genów kodujących białka. Introny mają dość stałe położenie w genach, które mało zmienia się w toku ewolucji. Tak więc geny, które kodują matrycowy RNAeukariotów, mają przerywaną budowę (ryc. 26.3).
Gen prokariotów znacznie różni się pod względem budowy od genu eukariotycznego. Po pierwsze, gen prokariotyczny nie posiada intronów. Przy tym cała kolejność genu ulega transkrypcji bezpośrednio do matrycowego RNA. Po drugie, jedna cząsteczka matrycowego RNA zwyczajnie koduje kilka białek. Odcinek DNA, kodujący tę mRNA nazywa się ope-ronem (ryc. 26.4). Przy tym wszystkie geny, z których zbudowany jest operon, mają tylko jeden promotor na początku operonu. Operon łączy kilku genów białek włączonych do wspólnego procesu i dlatego syntezowanych razem. Najbardziej znany jest laktozowy operon kodujący trzy białka, które biorą udział w spożyciu laktozy i operon tryptofany kodujący pięć białek, które uczestniczą w biosyntezie aminokwasu tryptofanu. Większość genów prokariotów połączonych jest w operony. Ciekawa rzecz to, że operony rzadko są spotykane u eukariotów.
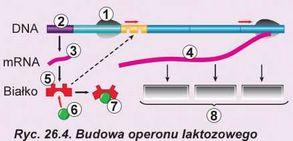
1. RNA-polimeraza. 2. Gen białka-regulatora. 3. mRNA bialka-regulatora. 4. mRNA trzech enzymów trawienia laktozy. 5. Białko-regulator. 6. Laktoza. 7. Nieaktywne bialko-regulator. 8. Enzymy trawienia laktozy. Zwróć uwagę, że białko-regulator może zablokować dostęp RNA-polimerazy do genów enzymów tylko w przypadku nieobecności laktozy w komórce.
Zastanów się
Wybierz jedną poprawną odpowiedź
1
Gen eukariotów zawiera informację o
AmRNA BmRNAIubtRNA CtRNAIubrRNA
D mRNA, tRNA lun rRNA E mRNA, tRNA, rRNA lub DNA
2
Gdy matrycowy RNA ma kolejność nukleotydów ACUGGCA, to łańcuch treściowy DNA miał kolejność
AACUGGCA В UGACCGU C ACGGTCA DACTGGCA E TGACCGT
3
Prawidłowe rozmieszczenie elementów genu jest następujące A start-kodon - promotor - część kodująca - stop-kodon - terminator В promotor - start-kodon - część kodująca - stop-kodon - terminator C stop-kodon - część kodująca - promotor - stop-kodon - terminator D promotor - start-kodon - część kodująca - terminator - stop-kodon E start-kodon - promotor - część kodująca - terminator - stop-kodon
4
W operon łączą się geny kodujące
A białka błony В białka niezbędne do biosyntezy białka
C różne RNA D białka niezbędne do jednego procesu E rRNA
5
Eukarioty wycinają introny podczas dojrzewania mRNA, dlatego że A rybosomowi niewygodnie jest pracować z długą mRNA В długie mRNA nie mogą opuścić jądra przez por jądrowy C introny zawierają informację o szkodliwych białkach D introny nie kodują części białek E introny przeszkadzają odczytywać egzony
Sformułuj odpowiedź w postaci kilku zdań
6
Dlaczego do syntezy mRNA wykorzystuje się antytreściowy łańcuszek DNA? Czy można odczytać ten sam gen z łańcuszka treściowego?
7
Długość genu zawsze jest większa od długości dojrzałej mRNA. Co jest tego przyczyną? Dokąd znikają zbyteczne fragmenty?
8
Dlaczego przy mieszaniu dojrzałej mRNA z łańcuchem antytreściowym DNA powstają pętle?
9
Czego w komórce jest więcej - genów czy różnych rodzajów białek syntezowanych według nich? Czy odpowiedź dotycząca eukariotów i prokariotów będzie jednakowa i dlaczego?
10
Czy ważne jest, w jakim kierunku będzie odczytywany gen podczas transkrypcji? Jak myślisz, co decyduje o kierunku odczytywania?
11
Co jest wspólnego między Terminatorem w wykonaniu Arnolda Schwarznegera w filmie o tym samym tytule i terminatorem genu?
Znajdź odpowiedź i postaraj się zrozumieć istotę problemu
12
Oceń wady i zalety operonowego systemu organizacji genów u prokariotów. Dlaczego eukarioty „odmówiły się” od takiego systemu?
13
Jaką rolę pełnią małe jądrowe RNA w procesie dojrzewania mRNA?
Dowiedz się samodzielnie i opowiedz innym
14
Oprócz genów kodujących enzymy w skład operonów wchodzą geny regulatorowe. Dokonaj opisu pracy takiej regulacji na przykładzie laktozowego operonu bakterii.
15
Alternatywne składanie genowe (splicing) znacznie rozszerza różnorodność białek organizmu. Na czym polega istota tego procesu i do jakich schorzeń może doprowadzić jego zakłócenie?
Dodatek VII
Hipoteza „jeden gen - jedno białko”
Przedtem uczeni sądzili, że każdemu białku odpowiada swój odcinek DNA, który nazywa się genem, i że jeden gen koduje jedno białko. Lecz już w końcu ubiegłego stulecia okazało się, że jeden gen może kodować kilku białek jednocześnie, a dla niektórych odcinków DNA końcowym produktem funkcjonalnym jest nawet nie białko, a cząsteczka RNA.
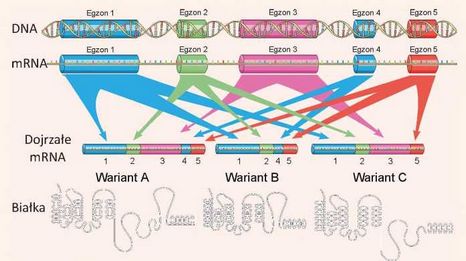
Rye. VII.1. Schemat alternatywnego składania genowego z pięcioma egzonami
W toku transkrypcji syntezuje się łańcuch RNA, zawierający introny i wszystkie pięć egzonów, lecz do dojrzałego mRNA docierają tylko niektóre egzony, więc w wyniku tego powstają trzy warianty mRNA, które odpowiadają trzem formom białkowego produktu.
Cząsteczka RNA otrzymana w toku transkrypcji DNA nie od razu uczestniczy w translacji i syntezie polipeptydowego łańcucha nowego białka, a najpierw doznaje znacznej reorganizacji - dojrzewania. Dopiero co syntezowana podczas transkrypcji cząsteczka RNA - transkrypt - zawiera w sobie egzony i introny. W toku dojrzewania cząsteczki RNA, oprócz wszyst-
kiego innego, zachodzi jej składanie (splicing): introny wycinają się, a egzony łączą się razem i taki już dojrzały matrycowy RNA (mRNA) jest podstawą do translacji. Za proces składania RNA w komórce odpowiada kompleks białek i cząsteczek RNA - spliceosom.
Lecz nie zawsze wszystkie egzony zszywają się i dostają do mRNA. Niektóre z nich w określonych warunkach w ciągu składania wycinają się wraz z intronami, a czasem niektóre introny nie wycinają się i zostają w mRNA. Wskutek tego zjawiska, który nazywa się alternatywnym składaniem, zachodzi translacja różnych wariantów białka z tego samego genu. Schematycznie proces alternatywnego składania podano na rycinie VII.1.
Składanie alternatywne było odkryte w końcu 70. lat XX wieku u adenowirusów, lecz jego rozpowszechnienie i znaczenie dla żywych organizmów udało się zrozumieć tylko niedawno.
Wychodząc z hipotezy, że jeden gen koduje jedno białko, uczeni nie mogli wyjaśnić, dlaczego człowiek ma tylko 23 tysiące genów, co nie jest o wiele więcej niż genów w genomie ob-leńca Caenorhabditis (około 20000), ale przy tym organizm ludzki jest bardziej skomplikowany niż obleńców. Analiza matrycowego RNA, przeprowadzona w roku 2008 przez uczonych z USA pokazała, że składaniu alternatywnemu poddaje się do 94% genów człowieka. Natomiast u Ca-enorhabditisa składanie alternatywne dotyczy tylko około 15% genów. Wynika z tego, że przy prawie takiej samej ilości genów w organizmie człowieka syntezuje się o wiele więcej różnych białek.
Najlepiej składanie alternatywne było poznane u drozofili, która w ciągu dziesięcioleci jest ulubionym obiektem różnorodnych badań genetycznych. Różnice między samicami i samcami u tych muszek owocowych wyznaczają kaskady wzajemnie powiązanych genów, które poddają się alternatywnemu składaniu. Przytoczmy przykład: płeć u drozofili determinowana jest przez ilość X-chromosomów, która wywiera wpływ na składanie alternatywne genu Sxi. Przy tym białko funkcjonalne Sxi odczytuje się tylko u samic. To białko reguluje alternatywne składanie genu Tra, wskutek jego działania zachodzi synteza funkcjonalnego białka Tra (tylko u samic). Z kolei w układzie nerwowym drozofili białko Tra reguluje alternatywne składanie genu Fru, ale już w taki sposób, że białko na otrzymanym mRNA u samic nie powstaje. Wskutek tego funkcjonalne białko Fru jest tylko u samców i właśnie ono wyznacza nawyki zachowawcze, niezbędne im do zalecania się do samic! W toku eksperymentu udowodniono, że sa-mice-mutanty z „męskim” wariantem białka Fru też zalecają się do samic.
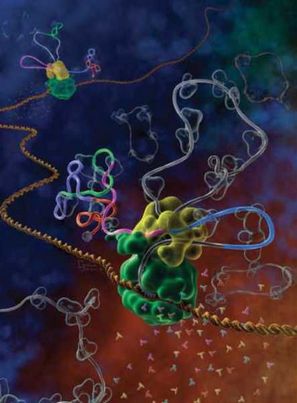
Rye. VII.2. Składanie alternatywne oczyma malarzy
Dwie RNA-polimerazy (zielone białko globularne) porusza się podwójną helisą DNA (pomarańczowego koloru) w głąb ryciny, syntezując cząsteczkę mRNA - traskrypt (szare odcinki wstążki). Syntezowany transkrypt RNA zawiera różne egzony (odcinki kolorowe) i introny (odcinki szare wstążki). Spliceosom (żółta globularna struktura) wycina kolejny intron (szara pętla ku górze) i łączy końce błękitnego i różowego egzonu. W już opracowany łańcuch RNA (po lewej stronie od spliceosomu) włączony jest zielony egzon. A drugi spliceosom (w głębi ryciny), na odwrót, wycina zielony egzon ze swego łańcucha RNA. Wycięte introny, zamknięte w pierścienie, płyną precz od spliceosomu.
Źródło: Biologia podręcznika dla klasy 9 Szałamow